Now, we have another two individuals with haplogroup J2 (highlighted in yellow):
Information about Kurdish autosomal DNA has been updated:
HarappaWorld
Dodecad K12b
Eurogenes K12b
McDonald
1x E1b1b1c1a (Alevi Kurmanji from Dersim/Turkey)
1x G2a (Alevi Kurmanji from Turkey)
2x J1 (Feyli, originally from Iran)
1x J1c3 (Sorani from Iran)
1x J2 (Zaza from Dersim/Turkey)
1x J2 (Kurmanji from Dohuk)
1x J2 (Kurmanji from Turkey)
1x J2a3a (J2a1a at 23andme; J2a4a at ISOGG 2009; he is M47+, M322+)(Yezidi from Iraq)
1x T (Sorani from Koysinjaq/Iraq)
1x R2a (Sorani from Sulaymaniyah/Iraq)
1x R1b1a2* (Kurmanji from Zakho/Iraq)
1x R1b1b2a (Zaza from Turkey)
1x R1b1 (P25+)(Kurmanji from Maras/Elbistan/Turkey)
1x R1a1a (Z93+, L342+, L657-)(Sorani from Sulaymaniyah/Iraq)
1x R1a1a (Z283+, Z282+, Z284-, M458-, Z280-, subclade 3 only his paternal great-grandfather is Kurdish from Turkey)
1x R1a1a (Alevi Zaza from Dersim/Turkey)
1x R1a1a (Alevi Kurmanji from Dersim/Turkey)
1x I2a2a* (old I2b1*; L1229-, L1230-, L1226-, L699-, L701-, L702-, L703-, L704-, M379) (Sorani from Sulaymaniyah/Iraq)
So far all tested SNPs of the I2a2a* individual turned out be negative.
More data can be found here:
Kurdish Y-DNA Part I
Kurdish Y-DNA Part II
Kurdish Y-DNA Part III
Kurdish Y-DNA Part IV
Kurdish Y-DNA Part V
Kurdish Y-DNA Part VI
Kurdish Y-DNA Part VIII
Thursday, June 28, 2012
mtDNA of Kurds V
Just an update (new entries are highlighted in yellow):
1x C4b (Alevi Kurmanji)
1x G2a (Sorani)
1x H5a1 (Sorani)
1x H13a2 (Alevi Kurmanji from Dersim)
1x H14 (Yezidi)
1x H15a1 (Sorani; mtDNA fully sequenced here and here)
1x H15b (Sorani)
1x HV (Sorani)
1x HV (Kurmanji from Zakho)
1x I5a (Zaza from Dersim)
1x J1b (Sorani)
1x J1c (Alevi Kurmanji from Dersim)
1x J2a1a (Kurd from Turkey)
1x N1b1 (Alevi Kurmanji from Dersim)
1x U1a1 (Zaza)
1x U1a1 (Sorani)
1x U5a1 (Kurmanji from Dohuk)
1x U8b (Feyli)
Some information about the latest entries:
mtDNA G2a:
The mtDNA haplogroup G2a is not known in populations from the Middle East but in Ainu from Japan and in Northeastern Siberia close to Bering Strait. There is only one reported case of a fully sequenced mtDNA G2a from the Caucasus (Georgian). However, this Georgian individual is G2a1b (not the same subbranch). Another curiosity is that this Kurdish individual has the 16172C mutation. 16172C was only described once in a Chinese individual. Strange enough, the Chinese individual does not belong to G2a but to the neighbor haplogroup G2b (to be precise G2b1a). My guess is that
a) it is a very, very rare coincidence to have the same mutation as two indepedent events,
b) 23andme is not testing for G2b mutations, or
c) the mtDNA G haplogroup tree needs to be updated.
Behar et al. 2012 estimated an age of 26788 ± 4618 years for G2.
Behar et al. 2012 estimated an age of 17146 ± 5270 years for G2a.
Here is a map of 23andme users with mtDNA G2a created by Evon_Evon who has a general interest in this mtDNA haplogroup.
mtDNA I5a:
The mtDNA Haplogroup I is found in Europe, Middle East and South Asia. Quintana-Murci et al., 2004, analyzed the mtDNA of 20 Kurds from Iran, and found one individual with the mtDNA I (1/20=5.0%). Several cases of I5a (Germany, Italy, Romania, Russia, USA) are described in the FTDNA mtDNA Haplogroup I Project.
Behar et al. 2012 estimated an age of 15116 ± 4128 years for I5a. Fully sequenced I5a mtDNA is available from Yemen, Dubai, and Turkey (GenBank). The individual from Turkey is the closest match for the Zaza (both are lacking the "Arabian mutations "G3705A","T5096C", and "G5773A").
mtDNA U5a1:
The mtDNA Haplogroup U5a1 is quiet common in Europe but it is also present in other parts of Eurasia being one of the most common and best described mtDNA haplogroups. Full mtDNA sepuences of the root of U5a1 are available from England, Spain, Caucasus, and Czech Republic at GenBank. The mtDNA of famous Cheddar Man from England (who lived 9,000 years ago) turned out be U5.
1x C4b (Alevi Kurmanji)
1x G2a (Sorani)
1x H5a1 (Sorani)
1x H13a2 (Alevi Kurmanji from Dersim)
1x H14 (Yezidi)
1x H15a1 (Sorani; mtDNA fully sequenced here and here)
1x H15b (Sorani)
1x HV (Sorani)
1x HV (Kurmanji from Zakho)
1x I5a (Zaza from Dersim)
1x J1b (Sorani)
1x J1c (Alevi Kurmanji from Dersim)
1x J2a1a (Kurd from Turkey)
1x N1b1 (Alevi Kurmanji from Dersim)
1x U1a1 (Zaza)
1x U1a1 (Sorani)
1x U5a1 (Kurmanji from Dohuk)
1x U8b (Feyli)
Some information about the latest entries:
mtDNA G2a:
The mtDNA haplogroup G2a is not known in populations from the Middle East but in Ainu from Japan and in Northeastern Siberia close to Bering Strait. There is only one reported case of a fully sequenced mtDNA G2a from the Caucasus (Georgian). However, this Georgian individual is G2a1b (not the same subbranch). Another curiosity is that this Kurdish individual has the 16172C mutation. 16172C was only described once in a Chinese individual. Strange enough, the Chinese individual does not belong to G2a but to the neighbor haplogroup G2b (to be precise G2b1a). My guess is that
a) it is a very, very rare coincidence to have the same mutation as two indepedent events,
b) 23andme is not testing for G2b mutations, or
c) the mtDNA G haplogroup tree needs to be updated.
Behar et al. 2012 estimated an age of 26788 ± 4618 years for G2.
Behar et al. 2012 estimated an age of 17146 ± 5270 years for G2a.
Here is a map of 23andme users with mtDNA G2a created by Evon_Evon who has a general interest in this mtDNA haplogroup.
mtDNA I5a:
The mtDNA Haplogroup I is found in Europe, Middle East and South Asia. Quintana-Murci et al., 2004, analyzed the mtDNA of 20 Kurds from Iran, and found one individual with the mtDNA I (1/20=5.0%). Several cases of I5a (Germany, Italy, Romania, Russia, USA) are described in the FTDNA mtDNA Haplogroup I Project.
Behar et al. 2012 estimated an age of 15116 ± 4128 years for I5a. Fully sequenced I5a mtDNA is available from Yemen, Dubai, and Turkey (GenBank). The individual from Turkey is the closest match for the Zaza (both are lacking the "Arabian mutations "G3705A","T5096C", and "G5773A").
mtDNA U5a1:
The mtDNA Haplogroup U5a1 is quiet common in Europe but it is also present in other parts of Eurasia being one of the most common and best described mtDNA haplogroups. Full mtDNA sepuences of the root of U5a1 are available from England, Spain, Caucasus, and Czech Republic at GenBank. The mtDNA of famous Cheddar Man from England (who lived 9,000 years ago) turned out be U5.
Thursday, June 21, 2012
Lactose Tolerance
The origin of domestic livestock (Goat, sheep, pig, cattle) started in the Middle East at least 10,000 years ago and spread from there all over the world. In the scientific publication “Domestication and early agriculture in the Mediterranean Basin: Origins, diffusion, and impact,” by Zeder et al., 2008, a comprehensive map is shown highlighting the historical importance of this region for mankind and its current diet.

The change of the diet from hunter/gatherers to farmers impacted not only the genetics of crop and livestock but also the genetics of humans. The most known and best described change is the ability of digesting and thus tolerating of milk as a food product, not only as infant but also as adult. Lactose, the sugar of milk, is the critical ingredient in milk that defines if someone can digest and thus tolerate milk or not. The responsible enzyme for digestion of lactose is called lactase, an enzyme that is highly expressed at early age in the small intestine and down-regulated after weaning, that's why adult hunter/gatherers are intolerant towards milk (lactose intolerant), while farmer populations can tolerate milk (lactose tolerant).
Dairy products (milk, yogurt, cheese) play an important role in the diet of the Iranian and Kurdish people, probably for thousands of years. It should be noted that Kurdish dairy products mostly rely on sheep and goat milk, while in Arabia camels were the main source of milk. In Europe bovine milk is dominating. Due to the size of camels and cattle it is very likely that Bedouins and Europeans consumed more dairy products than e.g. Iranians.
However, for quiet a while I am surprised to see how many Kurds are genetically lactose intolerant but physically lactose tolerant, even at high age. The best known SNP that is responsible for lactose tolerance is rs4988235.
1. People with "CC" at rs4988235 (also called C/T-13910) are genetically lactose intolerant; at 23andme it is "GG" (because MCM6 gene is in minus orientation but 23andme does not address it).
2. People with "CT" or "TT" at rs4988235 (also called C/T-13910) are genetically lactose tolerant; at 23andme it is "AG" or "AA" (because MCM6 gene is in minus orientation but 23andme does not address it)
Note: The actual gene for the enzyme lactase is called LCT and it is close to the gene MCM6. The MCM6 gene region seems to be important for the gene regulation of the LCT gene.
Let's take a look at the worldwide frequencies of rs4988235 (also called C/T-13910) in the Alfred database. The yellow bar shows the frequency of people that are genetically lactose intolerant (based on this one SNP), the green bar shows the frequency of people that are lactose tolerant:




Only 2-6% of Arabs, Bedouins, Druze, and Palestinians are genetically lactose tolerant.
Unfortunately, Northern Middle Eastern populations are not included in the ALFRED database but just looking at available data frequency of several Middle Eastern populations for this SNP rs4988235 it becomes obvious that rs4988235 cannot be only SNP responsible for lactose tolerance, there must be other independent mutations in the Middle Eastern genepool resulting in lactose tolerance.
Yuval Itan presented a nice map in his dissertation regarding the worldwide frequency of C-13910:

So, where can we find more data about other SNPs that are related to lactose tolerance in the Middle East?
Enattah et al., 2008, showed that other Middle Eastern populations are lactose tolerant without having the mutation at SNP rs4988235 (also called C/T-13910). Instead they found mutations very close to rs4988235 (all in intron13) and one on exon17 of the MCM6 gene. The latter one on exon17 (T/C-3712) seems to be Arab specific.


The allele frequency of these SNPs in the Middle East (from Enattah et al., 2008):

Based on these data it seems that the major SNPs for Saudi-Arabian lactose tolerance are found:
1. It is T/G-13915 + T/C-3712, both SNPs always occur in pairs, which shows the strong positive selection for lactose tolerance and the common origin of both SNPs.
2. Even though 23andme is just determining T/G-13915 but not T/C-3712, it's okay because the two SNPs always come as pairs.
So, if you are an Arab (especially Saudi-Arabian) and lactose tolerant you most likely have at least one C allele at T/C-3712 and one G at T/G-13915.
The latter one is covered by 23andme, look up rs41380347 (Note: Again, because MCM6 gene is in the minus orientation and 23andme does not address it, all nucleotides are switched in the following manner: "G=>C", "C=>G", "T=>A" and "A=>T"):
rs41380347: If you have "AA" then you don't have the "Arabian lactose tolerance".
rs41380347: If you have "AC" or even "CC", then you do have the "Arabian lactose tolerance".
Finally, let's take a look at the worldwide frequencies of rs41380347 (also called T/G-13915) in the Alfred database. The yellow bar shows the frequency of people that are genetically lactose intolerant (based on this "Arabian" SNP), the green bar shows the frequency of people that are lactose tolerant:

Yuval Itan presented a nice map in his dissertation regarding the worldwide frequency of T/G-13915 and two more African SNPs known to cause lactose persistence:

Here is another map summarizing all known lactose persistence SNPs and their worldwide frequencies:

Contrary, here is a map showing lactose persistence phenotype frequencies:

Summary:
Not all lactose tolerance SNPs are known, several SNPs resulting in lactose tolerance still need to be discovered.
One way to discover the relevant regions in the human genome for such SNPs in the Middle Eastern genepool is by family studies:
If some of your family members are lactose tolerant while other family members are lactose intolerant and if you have enough 23andme results of both groups within your family, you might be able to pinpoint the genomic region by exclusion of not relevant genomic regions.

The change of the diet from hunter/gatherers to farmers impacted not only the genetics of crop and livestock but also the genetics of humans. The most known and best described change is the ability of digesting and thus tolerating of milk as a food product, not only as infant but also as adult. Lactose, the sugar of milk, is the critical ingredient in milk that defines if someone can digest and thus tolerate milk or not. The responsible enzyme for digestion of lactose is called lactase, an enzyme that is highly expressed at early age in the small intestine and down-regulated after weaning, that's why adult hunter/gatherers are intolerant towards milk (lactose intolerant), while farmer populations can tolerate milk (lactose tolerant).
Dairy products (milk, yogurt, cheese) play an important role in the diet of the Iranian and Kurdish people, probably for thousands of years. It should be noted that Kurdish dairy products mostly rely on sheep and goat milk, while in Arabia camels were the main source of milk. In Europe bovine milk is dominating. Due to the size of camels and cattle it is very likely that Bedouins and Europeans consumed more dairy products than e.g. Iranians.
However, for quiet a while I am surprised to see how many Kurds are genetically lactose intolerant but physically lactose tolerant, even at high age. The best known SNP that is responsible for lactose tolerance is rs4988235.
1. People with "CC" at rs4988235 (also called C/T-13910) are genetically lactose intolerant; at 23andme it is "GG" (because MCM6 gene is in minus orientation but 23andme does not address it).
2. People with "CT" or "TT" at rs4988235 (also called C/T-13910) are genetically lactose tolerant; at 23andme it is "AG" or "AA" (because MCM6 gene is in minus orientation but 23andme does not address it)
Note: The actual gene for the enzyme lactase is called LCT and it is close to the gene MCM6. The MCM6 gene region seems to be important for the gene regulation of the LCT gene.
Let's take a look at the worldwide frequencies of rs4988235 (also called C/T-13910) in the Alfred database. The yellow bar shows the frequency of people that are genetically lactose intolerant (based on this one SNP), the green bar shows the frequency of people that are lactose tolerant:




Only 2-6% of Arabs, Bedouins, Druze, and Palestinians are genetically lactose tolerant.
Unfortunately, Northern Middle Eastern populations are not included in the ALFRED database but just looking at available data frequency of several Middle Eastern populations for this SNP rs4988235 it becomes obvious that rs4988235 cannot be only SNP responsible for lactose tolerance, there must be other independent mutations in the Middle Eastern genepool resulting in lactose tolerance.
Yuval Itan presented a nice map in his dissertation regarding the worldwide frequency of C-13910:

So, where can we find more data about other SNPs that are related to lactose tolerance in the Middle East?
Enattah et al., 2008, showed that other Middle Eastern populations are lactose tolerant without having the mutation at SNP rs4988235 (also called C/T-13910). Instead they found mutations very close to rs4988235 (all in intron13) and one on exon17 of the MCM6 gene. The latter one on exon17 (T/C-3712) seems to be Arab specific.

The allele frequency of these SNPs in the Middle East (from Enattah et al., 2008):

Based on these data it seems that the major SNPs for Saudi-Arabian lactose tolerance are found:
1. It is T/G-13915 + T/C-3712, both SNPs always occur in pairs, which shows the strong positive selection for lactose tolerance and the common origin of both SNPs.
2. Even though 23andme is just determining T/G-13915 but not T/C-3712, it's okay because the two SNPs always come as pairs.
So, if you are an Arab (especially Saudi-Arabian) and lactose tolerant you most likely have at least one C allele at T/C-3712 and one G at T/G-13915.
The latter one is covered by 23andme, look up rs41380347 (Note: Again, because MCM6 gene is in the minus orientation and 23andme does not address it, all nucleotides are switched in the following manner: "G=>C", "C=>G", "T=>A" and "A=>T"):
rs41380347: If you have "AA" then you don't have the "Arabian lactose tolerance".
rs41380347: If you have "AC" or even "CC", then you do have the "Arabian lactose tolerance".
Finally, let's take a look at the worldwide frequencies of rs41380347 (also called T/G-13915) in the Alfred database. The yellow bar shows the frequency of people that are genetically lactose intolerant (based on this "Arabian" SNP), the green bar shows the frequency of people that are lactose tolerant:

Yuval Itan presented a nice map in his dissertation regarding the worldwide frequency of T/G-13915 and two more African SNPs known to cause lactose persistence:

Here is another map summarizing all known lactose persistence SNPs and their worldwide frequencies:

Contrary, here is a map showing lactose persistence phenotype frequencies:

Summary:
Not all lactose tolerance SNPs are known, several SNPs resulting in lactose tolerance still need to be discovered.
One way to discover the relevant regions in the human genome for such SNPs in the Middle Eastern genepool is by family studies:
If some of your family members are lactose tolerant while other family members are lactose intolerant and if you have enough 23andme results of both groups within your family, you might be able to pinpoint the genomic region by exclusion of not relevant genomic regions.
Wednesday, June 20, 2012
Musical language groups
A new scientific article tries to answer the question if there is a correlation between ancestry and folk music. Based on the article, the answer is yes, there is a correlation between ancestry and folk music, to be more precisely maternal ancestry. The mothers are the carriers of folk songs.
Fortunately, Kurdish folk music was included in this study (total of 31 tested Eurasian nations). Kurdish folk music is a mayor part of Kurdish identity. The structure of Kurdish folk music seems to have similarities with Azeri, Warmia (Poland), Russian, Balkan, Bulgarian, and Komi folk music (see below).

A comparative phylogenetic study of genetics and folk music
Horolma Pamjav, Zoltán Juhász, Andrea Zalán, Endre Németh and Bayarlkhagva Damdin
Abstract:
Computer-aided comparison of folk music from different nations is one of the newest research areas. We were intrigued to have identified some important similarities between phylogenetic studies and modern folk music. First of all, both of them use similar concepts and representation tools such as multidimensional scaling for modelling relationship between populations. This gave us the idea to investigate whether these connections are merely accidental or if they mirror population migrations from the past. We raised the question; does the complex structure of musical connections display a clear picture and can this system be interpreted by the genetic analysis? This study is the first to systematically investigate the incidental genetic background of the folk music context between different populations. Paternal (42 populations) and maternal lineages (56 populations) were compared based on Fst genetic distances of the Y chromosomal and mtDNA haplogroup frequencies. To test this hypothesis, the corresponding musical cultures were also compared using an automatic overlap analysis of parallel melody styles for 31 Eurasian nations. We found that close musical relations of populations indicate close genetic distances (<0.05) with a probability of 82%. It was observed that there is a significant correlation between population genetics and folk music; maternal lineages have a more important role in folk music traditions than paternal lineages. Furthermore, the combination of these disciplines establishing a new interdisciplinary research field of “music-genetics” can be an efficient tool to get a more comprehensive picture on the complex behaviour of populations in prehistoric time.
Fortunately, Kurdish folk music was included in this study (total of 31 tested Eurasian nations). Kurdish folk music is a mayor part of Kurdish identity. The structure of Kurdish folk music seems to have similarities with Azeri, Warmia (Poland), Russian, Balkan, Bulgarian, and Komi folk music (see below).

A comparative phylogenetic study of genetics and folk music
Horolma Pamjav, Zoltán Juhász, Andrea Zalán, Endre Németh and Bayarlkhagva Damdin
Abstract:
Computer-aided comparison of folk music from different nations is one of the newest research areas. We were intrigued to have identified some important similarities between phylogenetic studies and modern folk music. First of all, both of them use similar concepts and representation tools such as multidimensional scaling for modelling relationship between populations. This gave us the idea to investigate whether these connections are merely accidental or if they mirror population migrations from the past. We raised the question; does the complex structure of musical connections display a clear picture and can this system be interpreted by the genetic analysis? This study is the first to systematically investigate the incidental genetic background of the folk music context between different populations. Paternal (42 populations) and maternal lineages (56 populations) were compared based on Fst genetic distances of the Y chromosomal and mtDNA haplogroup frequencies. To test this hypothesis, the corresponding musical cultures were also compared using an automatic overlap analysis of parallel melody styles for 31 Eurasian nations. We found that close musical relations of populations indicate close genetic distances (<0.05) with a probability of 82%. It was observed that there is a significant correlation between population genetics and folk music; maternal lineages have a more important role in folk music traditions than paternal lineages. Furthermore, the combination of these disciplines establishing a new interdisciplinary research field of “music-genetics” can be an efficient tool to get a more comprehensive picture on the complex behaviour of populations in prehistoric time.
Sunday, June 10, 2012
Kurdish Y chromosomes of a Central Anatolian village
Today, I want to focus on Omer Gokcumen's publication that I presented earlier.
He analyzed the Y-STR values of 4 Central Anatolian villages that are
in close proximity to each other, however, they are dominated by
different ethnic groups. For some reasons (that I don't want to discuss)
he used pseudonyms for the 4 villages, so please do not try to find the
locations on a map. One of 4 villages is described as Kurdish, Gokcumen
called this villages 'Dogukoy' in his publication.
Based on the figure 6 of the publication (see below), the following Y-STR haplogroups were observed in the Kurdish village of 'Dokukoy': E3b, I1b1, I1b2, J1, J2a, R1a, and R1b in a total of 31 tested individuals from 'Dokukoy'. E1b and J2a are dominating in the Kurdish village, while R1b, R1a, I1b1, I1b2, and J1 are present but relatively low.

A closer look at the Y-STR values of 31 tested individuals from 'Dokukoy' (can be found in the supplementary information) reveals that a lot of cases of J2a are identical or have a very narrow star-like occurence, so maybe Gokcumen et al. just tested a lot of relatives (one clan). A even more extreme pattern can be seen in the data set of the village 'Gocmenkoy' (Residents of 'Gocmenkoy' identify themselves with the Afsar clan of the Oguz tribe). Thus, I will not use any of the calculated Y-haplogroup frequencies of this publication, they are misleading. Pictures are sometimes better than 1000 words:

Green, red, blue, and yellow indicate the 'Gocmenkoy', 'Dogukoy', 'Eskikoy', and 'Merkez' settlements, respectively.
What can be used from the publication are the variation of observed Y-STR values of the Kurds of 'Dogukoy'.
I will compare these observed Y-STR haplotypes with published data and databases, maybe these 'Kurdish haplotypes' are present in other populations as well. As a starter, just the Modal haplotype for haplogroup J2a in Kurds from 'Dogukoy' (includes samples D05, D13, D16, and D26):
Update:
Some good matches for Modal haplotype for haplogroup J2a in Kurds from 'Dogukoy' at ysearch:
(distance 4/16=25%): BDVZY(Eash from Stiffansburg/Oberlangenegg, Switzerland) J2 (tested)
(distance 5/16=31%): NSUYM (Milan from Italy?) J2 (tested)
(distance 5/16=31%): EU9WG (Rachkovsky from Lithuania) J2a (tested)
After searching through ysearch and some scientific articles I came to the conclusion that the Modal haplotype for haplogroup J2a in Kurds from 'Dogukoy' is J2a4 and mostly likely J2a4b (M67(xM92) = old J2f). The majority of Kurds from Dogukoy (16 out 31 = 52%) belong to this Central Anatolian"Kurdish" J2a4b cluster.
Indeed, J2a4b is a very interesting haplogroup. It is found in high frequencies in North-Eastern Caucasian people, primarily Nakh language speakers.
As a consequence I took a closer look at STR values from Caucasus people, presented in Balanovsky et al., 2011.
First the Phylogenitic STR-network of the Caucasus based on Balanovsky et al., 2011.

The closest matches for the Modal haplotype for haplogroup J2a in Kurds from 'Dogukoy' are within cluster α or close by the founder haplotype of cluster α, mostly Nakh people (Ingush, Chechen).
(distance 5/17=29%): Ingush J2a4b* M67(xM92) ING-1341 outside clusters
(distance 5/17=29%): Dargins J2a4b* M67(xM92) DAG-299 α
(distance 6/17=35%): Chechen (Chechnya) J2a4b* M67(xM92) Chech-1021 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1354 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1358 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1353 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1359 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1365 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1421 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1321 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1330 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1366 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1434 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1405 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1413 α
(distance 6/17=35%): Chechen (Dagestan) J2a4b* M67(xM92) DAG06-495 outside clusters
(distance 6/17=35%): Chechen (Ingushetia) J2a4b* M67(xM92) Chech-1263 outside clusters
(distance 6/17=35%): Ossets-Iron J2a4b* M67(xM92) OSE-487 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1325 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1420 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) Chech-1246 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) Chech-1185 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) Chech-1240 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) Chech-1242 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) Chech-1244 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1011 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1319 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1392 outside clusters
(distance 6/17=35%): Avar J2a4b* M67(xM92) DAG-529 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) Chech-1221 outside clusters
Balanovsky et al., 2011 wrote:
Wikipedia states that:
Based on the figure 6 of the publication (see below), the following Y-STR haplogroups were observed in the Kurdish village of 'Dokukoy': E3b, I1b1, I1b2, J1, J2a, R1a, and R1b in a total of 31 tested individuals from 'Dokukoy'. E1b and J2a are dominating in the Kurdish village, while R1b, R1a, I1b1, I1b2, and J1 are present but relatively low.

A closer look at the Y-STR values of 31 tested individuals from 'Dokukoy' (can be found in the supplementary information) reveals that a lot of cases of J2a are identical or have a very narrow star-like occurence, so maybe Gokcumen et al. just tested a lot of relatives (one clan). A even more extreme pattern can be seen in the data set of the village 'Gocmenkoy' (Residents of 'Gocmenkoy' identify themselves with the Afsar clan of the Oguz tribe). Thus, I will not use any of the calculated Y-haplogroup frequencies of this publication, they are misleading. Pictures are sometimes better than 1000 words:

What can be used from the publication are the variation of observed Y-STR values of the Kurds of 'Dogukoy'.
I will compare these observed Y-STR haplotypes with published data and databases, maybe these 'Kurdish haplotypes' are present in other populations as well. As a starter, just the Modal haplotype for haplogroup J2a in Kurds from 'Dogukoy' (includes samples D05, D13, D16, and D26):
| DYS 393 | DYS 390 | DYS 19 | DYS 391 | DYS 385a | DYS 385b | DYS 439 | DYS 389I | DYS 392 | DYS 389II | DYS 458 | DYS 437 | DYS 448 | DYS 456 | DYS 438 | H4 GATA | DYS 635 |
| 12 | 22 | 14 | 11 | 13 | 15 | 12 | 12 | 11 | 29 | 16 | 15 | 19 | 15 | 9 | 21 | 22 |
Update:
Some good matches for Modal haplotype for haplogroup J2a in Kurds from 'Dogukoy' at ysearch:
(distance 4/16=25%): BDVZY(Eash from Stiffansburg/Oberlangenegg, Switzerland) J2 (tested)
(distance 5/16=31%): NSUYM (Milan from Italy?) J2 (tested)
(distance 5/16=31%): EU9WG (Rachkovsky from Lithuania) J2a (tested)
After searching through ysearch and some scientific articles I came to the conclusion that the Modal haplotype for haplogroup J2a in Kurds from 'Dogukoy' is J2a4 and mostly likely J2a4b (M67(xM92) = old J2f). The majority of Kurds from Dogukoy (16 out 31 = 52%) belong to this Central Anatolian"Kurdish" J2a4b cluster.
Indeed, J2a4b is a very interesting haplogroup. It is found in high frequencies in North-Eastern Caucasian people, primarily Nakh language speakers.
As a consequence I took a closer look at STR values from Caucasus people, presented in Balanovsky et al., 2011.
First the Phylogenitic STR-network of the Caucasus based on Balanovsky et al., 2011.

The closest matches for the Modal haplotype for haplogroup J2a in Kurds from 'Dogukoy' are within cluster α or close by the founder haplotype of cluster α, mostly Nakh people (Ingush, Chechen).
(distance 5/17=29%): Ingush J2a4b* M67(xM92) ING-1341 outside clusters
(distance 5/17=29%): Dargins J2a4b* M67(xM92) DAG-299 α
(distance 6/17=35%): Chechen (Chechnya) J2a4b* M67(xM92) Chech-1021 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1354 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1358 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1353 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1359 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1365 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1421 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1321 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1330 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1366 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1434 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1405 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1413 α
(distance 6/17=35%): Chechen (Dagestan) J2a4b* M67(xM92) DAG06-495 outside clusters
(distance 6/17=35%): Chechen (Ingushetia) J2a4b* M67(xM92) Chech-1263 outside clusters
(distance 6/17=35%): Ossets-Iron J2a4b* M67(xM92) OSE-487 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1325 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1420 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) Chech-1246 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) Chech-1185 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) Chech-1240 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) Chech-1242 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) Chech-1244 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1011 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1319 α
(distance 6/17=35%): Ingush J2a4b* M67(xM92) ING-1392 outside clusters
(distance 6/17=35%): Avar J2a4b* M67(xM92) DAG-529 outside clusters
(distance 6/17=35%): Ingush J2a4b* M67(xM92) Chech-1221 outside clusters
Balanovsky et al., 2011 wrote:
Although occupying a boundary position between Europe and the Near East, all four major Caucasus haplogroups show signs of a Near Eastern rather than European origin (fig. 2, supplementary fig. 1, Supplementary Material online). These four haplogroups reach their maximum (worldwide) frequencies in the Caucasus (table 2, fig. 2). They are either shared with Near East populations (G2a3b1-P303 and J2a4b*-M67(xM92)) or have ancestral lineages present there (G2a1*-P16(xP18) and J1*- M267(xP58)). Typical European haplogroups are very rare (I2a-P37.2) or limited to specific populations (R1a1a-M198) in the Caucasus. This pattern suggests unidirectional gene flow from the Near East toward the Caucasus, which could have occurred during the initial Paleolithic settlement or the subsequent Neolithic spread of farming. Archaeological data do not indicate a Near Eastern influence on the Neolithic cultures in the North Caucasus (Bader and Tsereteli 1989; Bzhania 1996; Masson et al. 1982), whereas Neolithization in the Transcaucasus was part of a Neolithic expansion that per- haps paralleled those occurring in Europe (Balaresque et al. 2010) and North Africa (Arredi et al. 2004). However, the current genetic evidence does not allow us to distinguish between Paleolithic and Neolithic models in shaping the genetic landscape of the North Caucasus.
Wikipedia states that:
Many scholars, such as Johanna Nichols[30] and Bernice Wuethrich [1] hold that the Dzurdzuks [ancient Nakh people living in Durdzukia] were descended from extremely ancient migrations from the Fertile Crescent to the Caucasus, perhaps due to population or political pressures back in the Fertile Crescent. Others who believe the so-called "Urartian version", such as George Anchabadze and Amjad Jaimoukha, still hold that those original migrants contributed to both the genetic and cultural traits of the modern Ingush and Chechens, but that the primary ancestors were Nakh-speaking migrants from what became Northeastern Urartu.
Saturday, June 9, 2012
Kurdish Y-DNA Part VI
Just a new update (highlighted in yellow):
1x E1b1b1c1a (Alevi Kurmanji from Dersim/Turkey)
1x G2a (Alevi Kurmanji from Turkey)
2x J1 (Feyli, originally from Iran)
1x J1c3 (Sorani from Iran)
1x J2 (Kurmanji from Turkey)
1x J2a3a (J2a1a at 23andme; J2a4a at ISOGG 2009; he is M47+, M322+)(Yezidi from Iraq)
1x T (Sorani from Koysinjaq/Iraq)
1x R2a (Sorani from Sulaymaniyah/Iraq)
1x R1b1a2* (Kurmanji from Zakho/Iraq)
1x R1b1b2a (Zaza from Turkey)
1x R1b1 (P25+)(Kurmanji from Maras/Elbistan/Turkey)
1x R1a1a (Z93+, L342+, L657-)(Sorani from Sulaymaniyah/Iraq)
1x R1a1a (Z283+, Z282+, Z284-, M458-, Z280-, subclade 3 only his paternal great-grandfather is Kurdish from Turkey)
1x R1a1a (Alevi Zaza from Dersim/Turkey)
1x R1a1a (Alevi Kurmanji from Dersim/Turkey)
1x I2a2a* (old I2b1*; L1229-, L1230-) (Sorani from Sulaymaniyah/Iraq)
The mutations Z282 and Z283 in Y-haplogroup R1a1a seem to have pretty much the same age.
The "Roots group" of the haplogroup I2a2a* (old I2b1*) in Europe seem to be L1229+ (group "1.1") or L1230+(group "1.2"), at least in the FTDNA I2b1/M223 Y-CLAN STUDY. The Sorani Kurd (at the bottom of the list above) was part of this group but turned out to be L1229- and L1230-. He is now in group "1.6".
The tree presented in the results part of the I2b1/M223 Y-CLAN STUDY shows the details/relationships of the different groups of I2a2a*. The position of the mentioned Sorani Kurd is highlighted in bold.
1- M223* (Roots-X)
1.1- L1229* (Roots)
1.1.1- L812* (Roots Group 1a/446 = 8, 438 = 8)
1.1.1.1- L319 (Roots Group 1a/446 = 8, 438 = 8)
1.1.2- L1230* (Roots Group 2a/446 = 9, 531 = 11)
1.2- Z161
1.2.1- L801
1.2.1.1- Z76* (Cont2a, Cont2to1, Cont1-X)
1.2.1.1.1- Z78* (Cont1-XX)
1.2.1.1.1.1- L1198 (Cont1, Cont1a, Cont1b)
1.2.1.1.1.1.1- Z79 (Cont1c)
1.2.1.1.2- P95 (Cont2b)
1.2.1.1.3- L1201 (Cont2b)
1.2.2- L623, L147.4 (Cont2c)
1.3- P78 (Cont3a)
1.3.1- L484 (Cont3a)
1.4- M284/L1195
1.4.1- L1193* (Isles E)
1.4.1.2 L1194 (Isles E)
1.4.2- L126, L137, L369 (Isles Limbo, Isles Sc)
1.5- L1228 (I2b1-X)
1.5.1- L1227 (I2b1-X)
1.6- L699/L701/L702/L703 (I2b1-XX)
1.6.1- L704 (I2b1-XX)
1.6.1.1- L1226 (I2b1-XX)
Update: The tree of I2a2a*-M223 was changed again. The position of the mentioned Sorani Kurd is highlighted in bold. I have my doubts about his position in group 1.3.2.
1.- M223* (Roots-X)
1.1- L1229* (Roots)
1.1.1- L812* (Roots Group 1a/446 = 8, 438 = 8)
1.1.1.1- L319 (Roots Group 1a/446 = 8, 438 = 8)
1.1.2- L1230 (Roots Group 2a Section 1/446 = 9, 531 = 11)
1.2- Z161
1.2.1- L801
1.2.1.1- Z76* (Cont2a, Cont2to1, Cont1-X)
1.2.1.1.1- Z78* (Cont1-XX)
1.2.1.1.1.1- L1198 (Cont1, Cont1a, Cont1b)
1.2.1.1.1.1.1- Z79 (Cont1c)
1.2.1.1.2- P95 (Cont2b)
1.2.1.1.3- L1201 (Cont2b)
1.2.2- L623, L147.4 (Cont2c)
1.3- L701/L702
1.3.1- P78 (Cont3a)
1.3.1.1- L484 (Cont3a)
1.3.2- L699/L703 (I2b1-XX)
1.3.2.1- L704 (I2b1-XX1)
1.3.2.1.1- L1226 (I2b1-XX1)
1.4- M284/L1195
1.4.1- L1193* (Isles E)
1.4.1.2 L1194 (Isles E)
1.4.2- L126, L137, L369 (Isles Limbo, Isles Sc)
1.5- L1228 (I2b1-X)
1.5.1- L1227 (I2b1-X)
Update June 22nd, 2012:
All recommended SNPs for group 1.3.2 (see above) turned out to be negative for the Sorani Kurd. He is L1226-, L699-, L701-, L702-, L703-, L704-, so he does not belong to this 1.3.2 group, either. Previously, he turned out to be L1229- and L1230- as well. The M233 tree at FTDNA needs to be updated, he belongs to group "1.- M223* (Roots-X)".
All these tested SNPs were observed in European M233 individuals but not in Asia. He is now testing M379, a SNP that was observed once in a sample from Pakistan (but nowhere else). This has been suggested by the Group Administrator of the FTDNA I2b1/M223 Y-CLAN Project.
More data can be found here:
Kurdish Y-DNA Part I
Kurdish Y-DNA Part II
Kurdish Y-DNA Part III
Kurdish Y-DNA Part IV
Kurdish Y-DNA Part V
Kurdish Y-DNA Part VII
Kurdish Y-DNA Part VIII
1x E1b1b1c1a (Alevi Kurmanji from Dersim/Turkey)
1x G2a (Alevi Kurmanji from Turkey)
2x J1 (Feyli, originally from Iran)
1x J1c3 (Sorani from Iran)
1x J2 (Kurmanji from Turkey)
1x J2a3a (J2a1a at 23andme; J2a4a at ISOGG 2009; he is M47+, M322+)(Yezidi from Iraq)
1x T (Sorani from Koysinjaq/Iraq)
1x R2a (Sorani from Sulaymaniyah/Iraq)
1x R1b1a2* (Kurmanji from Zakho/Iraq)
1x R1b1b2a (Zaza from Turkey)
1x R1b1 (P25+)(Kurmanji from Maras/Elbistan/Turkey)
1x R1a1a (Z93+, L342+, L657-)(Sorani from Sulaymaniyah/Iraq)
1x R1a1a (Z283+, Z282+, Z284-, M458-, Z280-, subclade 3 only his paternal great-grandfather is Kurdish from Turkey)
1x R1a1a (Alevi Zaza from Dersim/Turkey)
1x R1a1a (Alevi Kurmanji from Dersim/Turkey)
1x I2a2a* (old I2b1*; L1229-, L1230-) (Sorani from Sulaymaniyah/Iraq)
The mutations Z282 and Z283 in Y-haplogroup R1a1a seem to have pretty much the same age.
The "Roots group" of the haplogroup I2a2a* (old I2b1*) in Europe seem to be L1229+ (group "1.1") or L1230+(group "1.2"), at least in the FTDNA I2b1/M223 Y-CLAN STUDY. The Sorani Kurd (at the bottom of the list above) was part of this group but turned out to be L1229- and L1230-. He is now in group "1.6".
The tree presented in the results part of the I2b1/M223 Y-CLAN STUDY shows the details/relationships of the different groups of I2a2a*. The position of the mentioned Sorani Kurd is highlighted in bold.
1- M223* (Roots-X)
1.1- L1229* (Roots)
1.1.1- L812* (Roots Group 1a/446 = 8, 438 = 8)
1.1.1.1- L319 (Roots Group 1a/446 = 8, 438 = 8)
1.1.2- L1230* (Roots Group 2a/446 = 9, 531 = 11)
1.2- Z161
1.2.1- L801
1.2.1.1- Z76* (Cont2a, Cont2to1, Cont1-X)
1.2.1.1.1- Z78* (Cont1-XX)
1.2.1.1.1.1- L1198 (Cont1, Cont1a, Cont1b)
1.2.1.1.1.1.1- Z79 (Cont1c)
1.2.1.1.2- P95 (Cont2b)
1.2.1.1.3- L1201 (Cont2b)
1.2.2- L623, L147.4 (Cont2c)
1.3- P78 (Cont3a)
1.3.1- L484 (Cont3a)
1.4- M284/L1195
1.4.1- L1193* (Isles E)
1.4.1.2 L1194 (Isles E)
1.4.2- L126, L137, L369 (Isles Limbo, Isles Sc)
1.5- L1228 (I2b1-X)
1.5.1- L1227 (I2b1-X)
1.6- L699/L701/L702/L703 (I2b1-XX)
1.6.1- L704 (I2b1-XX)
1.6.1.1- L1226 (I2b1-XX)
Update: The tree of I2a2a*-M223 was changed again. The position of the mentioned Sorani Kurd is highlighted in bold. I have my doubts about his position in group 1.3.2.
1.- M223* (Roots-X)
1.1- L1229* (Roots)
1.1.1- L812* (Roots Group 1a/446 = 8, 438 = 8)
1.1.1.1- L319 (Roots Group 1a/446 = 8, 438 = 8)
1.1.2- L1230 (Roots Group 2a Section 1/446 = 9, 531 = 11)
1.2- Z161
1.2.1- L801
1.2.1.1- Z76* (Cont2a, Cont2to1, Cont1-X)
1.2.1.1.1- Z78* (Cont1-XX)
1.2.1.1.1.1- L1198 (Cont1, Cont1a, Cont1b)
1.2.1.1.1.1.1- Z79 (Cont1c)
1.2.1.1.2- P95 (Cont2b)
1.2.1.1.3- L1201 (Cont2b)
1.2.2- L623, L147.4 (Cont2c)
1.3- L701/L702
1.3.1- P78 (Cont3a)
1.3.1.1- L484 (Cont3a)
1.3.2- L699/L703 (I2b1-XX)
1.3.2.1- L704 (I2b1-XX1)
1.3.2.1.1- L1226 (I2b1-XX1)
1.4- M284/L1195
1.4.1- L1193* (Isles E)
1.4.1.2 L1194 (Isles E)
1.4.2- L126, L137, L369 (Isles Limbo, Isles Sc)
1.5- L1228 (I2b1-X)
1.5.1- L1227 (I2b1-X)
Update June 22nd, 2012:
All recommended SNPs for group 1.3.2 (see above) turned out to be negative for the Sorani Kurd. He is L1226-, L699-, L701-, L702-, L703-, L704-, so he does not belong to this 1.3.2 group, either. Previously, he turned out to be L1229- and L1230- as well. The M233 tree at FTDNA needs to be updated, he belongs to group "1.- M223* (Roots-X)".
All these tested SNPs were observed in European M233 individuals but not in Asia. He is now testing M379, a SNP that was observed once in a sample from Pakistan (but nowhere else). This has been suggested by the Group Administrator of the FTDNA I2b1/M223 Y-CLAN Project.
More data can be found here:
Kurdish Y-DNA Part I
Kurdish Y-DNA Part II
Kurdish Y-DNA Part III
Kurdish Y-DNA Part IV
Kurdish Y-DNA Part V
Kurdish Y-DNA Part VII
Kurdish Y-DNA Part VIII
Friday, June 8, 2012
Kurdish autosomal DNA based Eurogenes K12b
Today, I want to present Kurdish results using Eurogenes K12b, presented on gedmatch. Thanks to all the contributors of
this post.
Color coding for all relevant components:

Zaza1 from Turkey (KD001):

Zaza2 from Dersim/Turkey (KD024):

Alevi Kurmanji1 from Dersim/Turkey (KD002):

Alevi Kurmanji2 from Turkey (KD005):

Kurmanji from Turkey (KD006):

Kurmanji from Zakho/Iraq (KD007):

Kurmanji from Dohuk/Iraq (KD023):

Yezidi from Iraq (KD008):

Sorani1 from Sulaymaniyah/Iraq (KD009):

Sorani2 from Sulaymaniyah/Iraq (KD010):

Sorani3 from Sulaymaniyah/Iraq (KD011):

Sorani4 from Sulaymaniyah and Darband/Iraq (KD012):

Sorani5 from Koysinjaq/Iraq (KD013):

Sorani6 from Sulaymaniyah/Iraq (KD014):

Color coding for all relevant components:
Zaza1 from Turkey (KD001):

Zaza2 from Dersim/Turkey (KD024):

Alevi Kurmanji1 from Dersim/Turkey (KD002):

Alevi Kurmanji2 from Turkey (KD005):

Kurmanji from Turkey (KD006):

Kurmanji from Zakho/Iraq (KD007):

Kurmanji from Dohuk/Iraq (KD023):

Yezidi from Iraq (KD008):

Sorani1 from Sulaymaniyah/Iraq (KD009):

Sorani2 from Sulaymaniyah/Iraq (KD010):

Sorani3 from Sulaymaniyah/Iraq (KD011):

Sorani4 from Sulaymaniyah and Darband/Iraq (KD012):

Sorani5 from Koysinjaq/Iraq (KD013):

Sorani6 from Sulaymaniyah/Iraq (KD014):

Kurdish autosomal DNA: How are we mixed?
I. So what can we learn from the presented HarappaWorld results?
Only the components that are higher than 1% and occur in all presented samples can be considered as 'Kurdish' or 'Iranian'. Components that only occur in some samples are more likely due to individual 'family history'.
This helps us excluding most components that are not 'Iranian', i.e.
SE-Asian,
Siberian,
NE-Asian,
Papuan,
American,
Beringian,
San,
E-African,
Pygmy, and
W-African.
The South-Asian component is a tricky one since it is completely missing in one individual (Sorani Kurd5), but present in low amounts (Average 2%) in the other individuals.
The typical admixture of KurdishDNA based on HarappaWorld consists of 5 components (average of all 14 individuals):
Caucasus (43%),
Baloch (26%),
SW-Asian (14%),
Mediterranean (7%),
NE-European (6%), and
Other (6%).
II. What can we learn from the presented Dodecad K12b results?
Only the components that are higher than 1% and occur in all presented samples can be considered as 'Kurdish' or 'Iranian'. Components that only occur in some samples are more likely due to individual 'family history'.
This helps us excluding most components that are not 'Iranian', i.e.
Siberian,
NW-African,
SE-Asian,
E-African,
E-Asian, and
Sub_Saharan
Again, the South-Asian component is a tricky one since it is completely missing in one individual (Sorani Kurd5), but present in low amounts (Average 2%) in the other individuals.
The typical admixture of KurdishDNA based on Dodecad K12b consists of 5 components (average of all 14 individuals):
Caucasus (41%),
Gedrosia (27%),
SW-Asian (14%),
Atlantic-Mediterranean (7%),
N-European (6%), and
Other (5%).
The outcome of both HarappaWorld and Dodecad K12b is the same.
Last question for today: Do we see any gradient of these 5 components within the 14 samples?
Yes, we do. Between samples from the North vs from the South. The Northern samples have more of the 3 components Caucasus, Atlantic-Mediterranean/Mediterranean and N-European/NE-European, while the Southern samples have more of the 2 components Gedrosia/Baloch and SW-Asian.
Only the components that are higher than 1% and occur in all presented samples can be considered as 'Kurdish' or 'Iranian'. Components that only occur in some samples are more likely due to individual 'family history'.
This helps us excluding most components that are not 'Iranian', i.e.
SE-Asian,
Siberian,
NE-Asian,
Papuan,
American,
Beringian,
San,
E-African,
Pygmy, and
W-African.
The South-Asian component is a tricky one since it is completely missing in one individual (Sorani Kurd5), but present in low amounts (Average 2%) in the other individuals.
The typical admixture of KurdishDNA based on HarappaWorld consists of 5 components (average of all 14 individuals):
Caucasus (43%),
Baloch (26%),
SW-Asian (14%),
Mediterranean (7%),
NE-European (6%), and
Other (6%).
II. What can we learn from the presented Dodecad K12b results?
Only the components that are higher than 1% and occur in all presented samples can be considered as 'Kurdish' or 'Iranian'. Components that only occur in some samples are more likely due to individual 'family history'.
This helps us excluding most components that are not 'Iranian', i.e.
Siberian,
NW-African,
SE-Asian,
E-African,
E-Asian, and
Sub_Saharan
Again, the South-Asian component is a tricky one since it is completely missing in one individual (Sorani Kurd5), but present in low amounts (Average 2%) in the other individuals.
The typical admixture of KurdishDNA based on Dodecad K12b consists of 5 components (average of all 14 individuals):
Caucasus (41%),
Gedrosia (27%),
SW-Asian (14%),
Atlantic-Mediterranean (7%),
N-European (6%), and
Other (5%).
The outcome of both HarappaWorld and Dodecad K12b is the same.
Last question for today: Do we see any gradient of these 5 components within the 14 samples?
Yes, we do. Between samples from the North vs from the South. The Northern samples have more of the 3 components Caucasus, Atlantic-Mediterranean/Mediterranean and N-European/NE-European, while the Southern samples have more of the 2 components Gedrosia/Baloch and SW-Asian.
Kurdish autosomal DNA based on Dodecad K12b
Today, I want to present Kurdish results using Dienekes's tool called Dodecad K12b, presented on gedmatch. Thanks to all the contributors of
this post.
Results were used to determine biogeographical ancestry:
View Genome mapping of Kurds by Palisto in a larger map
To see it in direct comparison McDonald's maps, click here.
Color coding for all relevant components of Dodecad K12b:

Zaza1 from Turkey (KD001):

Zaza2 from Dersim/Turkey (KD024):

Alevi Kurmanji1 from Dersim/Turkey (KD002):

Alevi Kurmanji2 from Turkey (KD005):

Kurmanji from Turkey (KD006):

Kurmanji from Zakho/Iraq (KD007):

Kurmanji from Dohuk/Iraq (KD023):
 Yezidi from Iraq (KD008):
Yezidi from Iraq (KD008):

Sorani1 from Sulaymaniyah/Iraq (KD009):

Sorani2 from Sulaymaniyah/Iraq (KD010):

Sorani3 from Sulaymaniyah/Iraq (KD011):
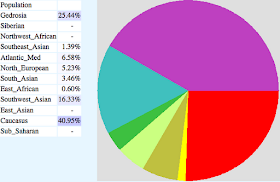
Sorani4 from Sulaymaniyah and Darband/Iraq (KD012):

Sorani5 from Koysinjaq/Iraq (KD013):

Sorani6 from Sulaymaniyah/Iraq (KD014):

Feyli (KD025):

Feyli (KD026):

Feyli (KD027):

Sorani from Sulaymaniyah/Iraq (KD032):

Sorani from Sulaymaniyah/Iraq (KD033):

Sorani from Sulaymaniyah/Iraq (KD034):

1/2Alevi Kurmanji-1/2Sunni Kurmanji from Bingöl (Zazaki: Çolig, Kurmanji: Çewlîg), Kighi, Turkey (KD036):

Results were used to determine biogeographical ancestry:
View Genome mapping of Kurds by Palisto in a larger map
To see it in direct comparison McDonald's maps, click here.
Color coding for all relevant components of Dodecad K12b:

Zaza1 from Turkey (KD001):

Zaza2 from Dersim/Turkey (KD024):

Alevi Kurmanji1 from Dersim/Turkey (KD002):

Alevi Kurmanji2 from Turkey (KD005):

Kurmanji from Turkey (KD006):

Kurmanji from Zakho/Iraq (KD007):

Kurmanji from Dohuk/Iraq (KD023):


Sorani1 from Sulaymaniyah/Iraq (KD009):

Sorani2 from Sulaymaniyah/Iraq (KD010):

Sorani3 from Sulaymaniyah/Iraq (KD011):

Sorani4 from Sulaymaniyah and Darband/Iraq (KD012):

Sorani5 from Koysinjaq/Iraq (KD013):

Sorani6 from Sulaymaniyah/Iraq (KD014):

Feyli (KD025):

Feyli (KD026):

Feyli (KD027):

Sorani from Sulaymaniyah/Iraq (KD032):

Sorani from Sulaymaniyah/Iraq (KD033):

Sorani from Sulaymaniyah/Iraq (KD034):

1/2Alevi Kurmanji-1/2Sunni Kurmanji from Bingöl (Zazaki: Çolig, Kurmanji: Çewlîg), Kighi, Turkey (KD036):
